![]()


Новости
- НОВОСТИ РОССИЙСКОГО ОФИСА СТХАПАТЬЯ ВЕДЫ
- НОВОСТИ МЕЖДУНАРОДНОГО ОФИСА СТХАПАТЬЯ ВЕДЫ
- ПРОГРЕССИВНЫЕ ИДЕИ И ТЕНДЕНЦИИ В СОВРЕМЕННОЙ АРХИТЕКТУРЕ
| Навигация | ||
| ^ к списку ^ | ||
 08.05.21
08.05.21
Паттерны и характеристики ЭЭГ при обращении лицом в кардинальных направлениях
Международный журнал психологических исследований; Том 13, № 2; 2021
ISSN 1918-7211 E-ISSN 1918-722X
Опубликовано Канадским центром науки и образования
Паттерны и характеристики ЭЭГ при обращении лицом в кардинальных направлениях
Фредерик Т. Трэвис 1, Джонатан Б. Липман 2, Ниязи Парим 1, Питер Л. Ходак 1 и Жаклин Дж. Лите 3
1 Центр мозга, сознания и познания, Международный Университет Махариши, Фэйрфилд, штат Айова, 52557, США
2 Институт Ведической Архитектуры Махариши, Международный университет Махариши, Фэрфилд, штат Айова, 52556, США
3 Отделение неврологии и поведения, Уэслианский Университет, Мидлтаун, Коннектикут 53562, США Для корреспонденции: Frederick Travis, 1000 North 4th Street, MR 683, Fairfield, IA 52557, Соединенные Штаты Америки.
Получено: 20 марта 2021 г.
Принято: 19 апреля 2021 г.
Интернет-публикация: 21 апреля 2021 г.
Введение
1) Предпосылки и цели: положение в пространстве и течение времени закодированы в возбуждении таламической, гиппокампальной и энторинальной коры у грызунов. Сообщалось о клетках направления головы у свободно движущихся обезьян, а у людей наблюдались разные мозговые паттерны во время игры в навигационную видеоигру и в ответ на изменения электромагнитных полей. Чувствительность организмов к окружающей среде и электромагнитным сигналам может объяснить рекомендации традиционной системы архитектуры, архитектуры Васту, которая рекомендует выравнивать дома по сторонам света.
2) Гипотеза: архитектура Васту предсказывает, что ориентация на восток и север более благоприятна, чем на запад и юг. Если ориентация на восток и север более предпочтительны, тогда испытуемые должны демонстрировать отчетливые паттерны ЭЭГ и улучшенные характеристики при обращении лицом на восток и север по сравнению с западом или югом.
3) Материалы и методы. Сравнивались паттерны когерентности ЭЭГ из 32-канальной ЭЭГ и время выполнения сборки пазлов, когда испытуемые смотрели в четырехсторонних направлениях.
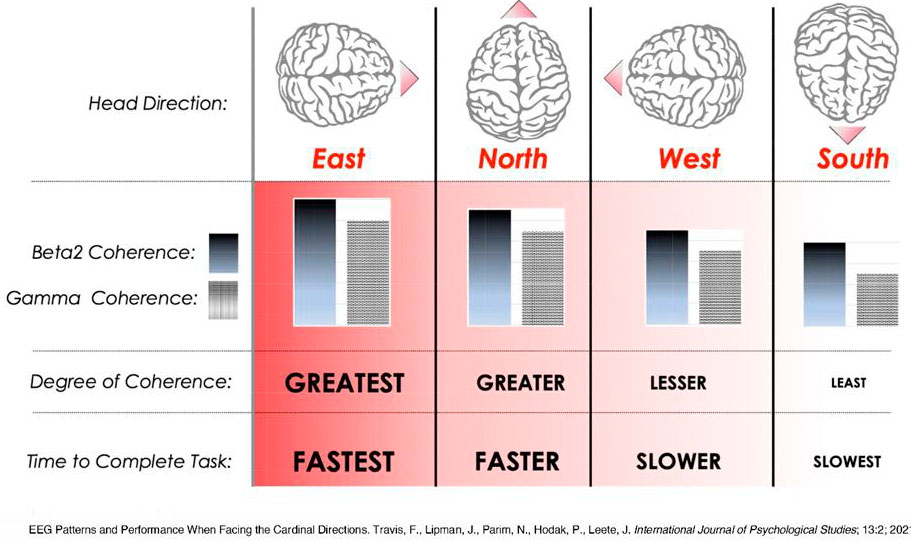
4) Результаты. Когда испытуемые смотрели на восток и север, фронтальная бета2- и гамма-когерентность ЭЭГ была значительно выше, и они собирали пазлы значительно быстрее, чем при взгляде на запад или юг.
5) Обсуждение: результаты мозга соответствуют данным о производительности. Лучшая фокусировка, которая разумно связана с более высокой производительностью, связана с более высокими уровнями бета 2- и гамма-когерентности.
6) Вывод: эти данные подтверждают возможность того, что человеческий мозг может быть чувствительным к сторонам света. Это подчеркивает, насколько тесно мы связаны с окружающей средой, и предлагает фактор, который может быть важным при ориентации рабочих пространств и проектировании классных комнат.
Ключевые слова: архитектура, стороны света, когерентность ЭЭГ, клетки направления головы, Ваасту.
1. Введение
Физическая среда, по-видимому, закодирована в паттернах возбуждения в таламических, гиппокампальных и энторинальных нейронах. О'Киф в 1971 году впервые сообщил об увеличении частоты возбуждения клеток гиппокампа, когда крысы перемещались в разные области своего лабиринта (O'Keefe & Dostrovsky, 1971). Работая с О'Кифом, Мэй-Бритт и Эдвард Мозер определили «сеточные» клетки в энторинальной коре головного мозга крысы, которые срабатывают, когда животное приближается к точкам регулярной сетки, охватывающей окружающую среду (Hafting et al., 2005; Moser et al., 2017). Эти трое были удостоены Нобелевской премии в 2014 году за свои открытия (Abbott & Callaway, 2014).
В недавней статье в Scientific American подробно описаны взаимосвязанные подкорковые сети, которые кодируют положение организма в пространстве и течение времени (Moser & Moser, 2016). Эти ментальные карты включают в себя: размещение клеток в гиппокампе, которые постоянно срабатывают, когда человек находится в определенном месте в знакомом пространственном поле (O'Keefe et al., 1975), ячейки сетки, которые обеспечивают нейронное представление пространства с сетями наверху энторинальной коры, отображающей более короткие расстояния, и те, что в нижней части коры, отображающие большие расстояния - до метров между точками сетки (Fyhn et al., 2008), пограничные клетки, которые отмечают основные границы в окружающей среде (Santos-Pata et al., 2017), и клетки направления головы, которые срабатывают максимально, когда организм смотрит в разные стороны (Butler et al., 2017; Taube, 1995). Эти сети были обнаружены у крыс, поскольку глубинные электроды можно было использовать для изучения глубоких структур мозга.
Клетки направления головы в предубикулуме, структуре, которая находится рядом с гиппокампом, были измерены у свободно движущихся обезьян. Скорость активации этих клеток была до 100 раз выше, когда обезьяны смотрели в одном направлении, чем когда они смотрели в другую сторону, с настройкой на 76 градусов (Butler et al., 2017). Эти клетки срабатывали, даже когда детали просмотра были затемнены или комната была затемнена, что свидетельствует о том, что эти клетки чувствительны к направлению, а не к визуальным стимулам.
Исследования показывают, что окружающая среда также может быть отображена в функционировании человеческого мозга. В одном исследовании сообщается о различных моделях возбуждения у нейрохирургических пациентов, играющих в навигационную видеоигру, когда они двигались по часовой стрелке, по сравнению с траекторией против часовой стрелки в видеоигре (Jacobs et al., 2010). Эти ячейки, по-видимому, кодировали информацию как о местоположении, так и о направлении. Другое исследование сообщает, что паттерны ЭЭГ в диапазоне 6-12 Гц были значительно уменьшены, когда ток передавался через три вложенных набора ортогональных катушек по сравнению с условиями отсутствия тока (Wang et al., 2019).
Более 5000 лет назад система архитектуры Ведической традиции, архитектура Васту, рекомендовала ориентировать дома и комнаты по сторонам света. Ориентация физической структуры может выровнять деятельность в здании по сторонам света. Архитектура Васту предсказывает, что ориентация на восток и север более благоприятна, чем на запад и юг (Lipman et al., 2021).
1.1 Гипотеза исследования
Если ориентация на восток и север более предпочтительны, тогда испытуемые должны демонстрировать разные паттерны ЭЭГ и иметь лучшие характеристики при обращении лицом на восток и север по сравнению с западом или югом.
2. Материалы и методы
2.1 Субъекты
Двадцать восемь испытуемых откликнулись на плакаты, рекламирующие исследование функционирования мозга при сборке пазлов. Их средний возраст составлял 50,0 ± 20,8 года, от 18 до 80 лет. Одиннадцать были женщинами. Все были психически и физически здоровы. Все они правши.
Испытуемые были набраны для составления пазлов, так, чтобы они не понимали гипотезы исследования. Это, похоже, сработало. Никто не спросил, тестируем ли мы мозговые паттерны и направление головы. Экспериментатор настойчиво нажимал на секундомер после каждого пазла и сообщал испытуемым, насколько они быстры.
2.2 Порядок проведения
Субъекты приходили в мозговой центр университета по отдельности ближе к вечеру. Они заполнили формы согласия, в то время как 32 датчика были применены в системе 10-10 с ориентирами лба и мочки левого и правого уха. Сопротивление каждого датчика было
Затем испытуемые переходили к столу в мозговом центре, который был выровнен по сторонам света. На каждом месте вокруг стола было сложено по четыре пазла. Каждый пазл мозаики состоял из 24 деталей.
Мы использовали квази-рандомизацию, чтобы равное количество испытуемых начинало смотреть в каждую из четырех сторон света, и чтобы их первый пазл случайным образом был одним из четырех. Таким образом, конкретное кардинальное направление не могло бы иметь одну и ту же загадку для всех субъектов.
Время завершения пазла было записано в десятых долях секунды. ЭЭГ регистрировалась с помощью системы BIOSEMI ActiveTwo ( www.BIOSEMI.COM ), когда испытуемые собирали каждый пазл. Закончив с одним пазлом, испытуемый вставал и переходил к следующему стулу, двигаясь по часовой стрелке. Все данные были оцифрованы в режиме онлайн со скоростью 256 точек в секунду, без высокочастотных или низкочастотных фильтров, и сохранены для последующего анализа с помощью Brain Vision Analyzer.
В результате этой процедуры семь испытуемых стали смотреть на восток, семь - на север и так далее. Пазлы по каждому направлению света также были рандомизированы. Дважды в первом направлении, с которого начинали испытуемые, был пазл 1, дважды пазл 2, дважды пазл 3 и один раз пазл 4.
2.2.1 Инструменты для тестирования: пазлы мозаики
Пазлы состояли из 24 частей и были рекомендованы для детей от 5 лет и старше. Мы использовали пазлы из 24 частей, а не более сложные пазлы, такие как пазлы из 100 частей, по трем причинам. Во-первых, для решения более сложных пазлов потребуется больше времени, что приведет к усталости при тестировании и более длительной записи. Во-вторых, усталость от тестирования может привести к реактивности испытуемых, и испытуемые могут быть менее склонны уделять все внимание сборке третьей и четвертой головоломок. В-третьих, усталость при тестировании может привести к большему количеству артефактов движения, которые могут загрязнить ЭЭГ.
2.3 Анализ данных: ЭЭГ
Данные ЭЭГ анализировали с помощью анализатора Brain Vision. Во время периодов сборки пазлов были выбраны три минуты необработанной ЭЭГ. Необработанная ЭЭГ была визуально проверена на предмет движения тела, скольжения электродов или глазных артефактов, помечена вручную и не включена в спектральный анализ. Необработанные данные были повторно привязаны к усредненным связанным ушам, подверглись цифровой фильтрации в полосовом фильтре 2,0 - 50 Гц со спадом 48 дБ и быстрому преобразованию Фурье за 2-секундные эпохи с использованием окна Хеннинга с началом и смещением 10%. . Когерентность рассчитывалась с шагом 0,5 Гц в шести частотных диапазонах - дельта (0-4 Гц), тета (4-8 Гц), альфа1 (8-10 Гц), альфа2 (10-12 Гц), бета (12-20 Гц), бета2 (20-30 Гц) и гамма (30-50 Гц). Паттерны когерентности ЭЭГ отражают функциональную связь (Thatcher et al., 1986), обмен информацией (Petsche et al., 1997) и функциональную координацию (Gevins et al., 1989) между областями мозга, что может отражать активность подкорковых областей. Чтобы уменьшить количество сравнений, оценки когерентности были усреднены по 36 парам когерентности среди девяти фронтальных датчиков (AF3, AF4, F3, F4, F7, F8, Fz, FC1, FC2), 36 возможных пар среди девяти теменных датчиков (PO3, PO4, P3, P4, P7, P8, Pz, CP1, CP2) и четыре пары когерентности между лобными и теменными датчиками (F3 / P3, F4 / P4, AF3 / P3, AF4 / P4).
2.4 Анализ данных: пазлы
Время завершения сборки пазлов фиксировалось секундомером с точностью до десятых долей секунды.
2.5 Статистический анализ
MANOVA с повторными измерениями использовался для проверки оценок согласованности с направлением в качестве повторного измерения и трех областей мозга (лобной, теменной или передне-теменной) и семи частот в качестве переменных (SPSS 13.0). Индивидуальные повторные измерения ANOVA проводились при наличии значительных эффектов взаимодействия.
3. Полученные результаты
3.1 Анализ согласованности во время завершения сборки пазла
Комплексные повторные измерения MANOVA с направлением в качестве повторного измерения и трех областей мозга и семи частот в качестве переменных дали значимые взаимодействия мозг x частота (F (12, 324) = 17,0, p <0,001). Таким образом, повторные измерения MANOVA проводились в каждой области мозга.
В то время как не было значительных основных эффектов или взаимодействий в теменной и передне - теменной когерентности, во фронтальной когерентности наблюдалось существенное частотное взаимодействие. (F (18,504) = 1,8, p = 0,020). Таким образом, индивидуальные повторные измерения ANOVA были проведены для фронтальной когерентности в каждой полосе частот. Были только значимые эффекты направления в бета2 и гамма-когерентности (бета2: F (3,84) = 4,0, p = 0,010; гамма: F (3,84) = 2,8, p = 0,045). Значимые эффекты были линейными: наибольшая когерентность при обращении лицом на восток, затем согласованность при взгляде на север, затем на запад и наименьшая при обращении лицом на юг (бета2: F (1,28) = 6,2, p = 0,019; гамма (F (1,28) = 5,1, p = 0,032) Эти различия когерентности видны на рисунке 1. Частоты Beta2 и гамма находятся слева от рисунка и обведены кружком.
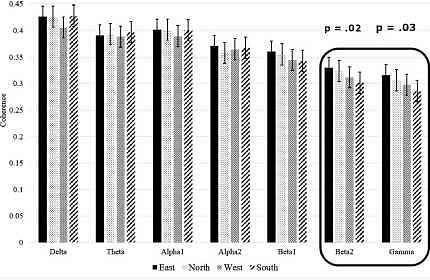
Рисунок 1. Фронтальная когерентность, усредненная по сторонам света.
Фронтальная когерентность при сборке пазлов была выше по частотам бета2 и гамма, если смотреть на восток или север, по сравнению с западом или югом.
3.2 Среднее время выполнения (сборки) пазлов
Время, необходимое для выполнения (сборки) пазлов, усреднялось в пределах каждого кардинального направления и сравнивалось с ANOVA с повторными измерениями. Этот анализ дал существенный основной эффект для направления скорости завершения сборки пазла (F (1,27) = 4,8, p = 0,037). Значительные эффекты были линейными: время завершения сборки пазла быстрее, если смотреть на восток, север, запад и юг. Эти результаты хорошо видны на Рисунке 2, где показано время, необходимое для выполнения сборки пазлов в четырех направлениях.
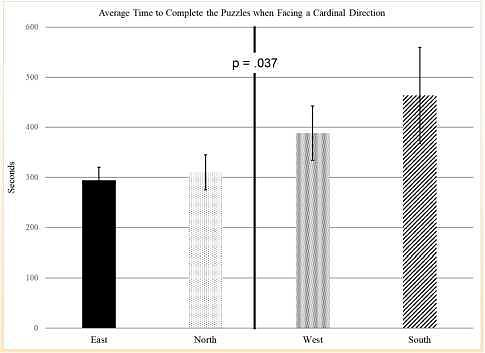
Рисунок 2. Время выполнения (сборки) пазлов.
Среднее время для решения четырех пазлов. Время обработки было быстрее, если смотреть на восток, затем на север, запад и юг.
4. Обсуждение
Фронтальная бета2- и гамма-когерентность ЭЭГ испытуемых была значительно выше, и они собирали пазлы значительно быстрее, когда смотрели на восток и север, чем на запад и юг. Результаты мозга соответствуют данным о производительности. Лучшая фокусировка, которая будет поддерживать более высокую производительность, связана с более высокими уровнями бета2- и гамма-когерентности (Jensen et al., 2007). Бета2- и гамма-когерентность возникает в результате локальной обработки в пределах короткодействующих связей, ответственных за распознавание объектов и целенаправленное поведение (Lubar, 1997; Singer, 1999). Таким образом, более высокая бета2- и гамма-когерентность наряду с более высокой производительностью при обращении к определенным сторонам света подтверждают возможность того, что человеческий мозг может быть чувствителен к сторонам света.
4.1 Ограничения исследования и будущие исследования
Проект был строгим: последовательность пазлов и начальное направление для первого пазла были рандомизированы, и испытуемые не знали гипотезы исследования. Однако количество испытуемых было небольшим (N = 28). В будущих исследованиях можно было бы воспроизвести эти результаты на более широкой популяции испытуемых и проверить влияние направления головы на реальные задачи, такие как способность запоминания учащихся, решение сложных задач или компьютерное кодирование.
5. Заключение
В этом слепом случайном исследовании направление головы было связано с более высокой скоростью сборки пазлов и более высокими уровнями бета2- и гамма-когерентности. Эти данные подтверждают возможность того, что человеческий мозг может быть чувствительным к сторонам света. Это подчеркивает, насколько тесно мы связаны с внешней средой.
Эти результаты могут послужить основой для дизайнерских решений и объяснить вариативность исследований в области образования - ориентация парт в классах может влиять на бдительность учащихся и, следовательно, на обучение, а также на исследования в области бизнеса - ориентация рабочих мест может влиять на производительность. Будущие исследования могут исследовать эти возможности.
Ссылки
Abbott, A., & Callaway, E. (2014). Prize for place cells. Nature, 514(7521), 153. doi.org/10.1038/514153a
Butler, W. N., Smith, K. S., van der Meer, M. A., & Taube, J. S. (2017). The Head-Direction Signal Plays a Functional Role as a Neural Compass during Navigation. Current Biology, 27(15), 2406. doi. org/10.1016/j. cub.2017.07.032
Fyhn, M., Hafting, T., Witter, M. P., Moser, E. I., & Moser, M. B. (2008). Grid cells in mice. Hippocampus, 18(12), 1230-1238. doi.org/10.1002/hipo.20472
Gevins, A. S., Bessler, S. L., Morgan, N. H., Cutillo, B. A., White, R. M., Greer, D. S., & Illes, J. (1989). Event- related covariances during a bimanual visuomotor task. I. methods and analysis of stimulus- and response-locked data. Electroencephalography & Clinical Neurophysiology, 74(1), 58-75. doi.org/10.1016/0168- 5597(89)90052-X
Hafting, T., Fyhn, M., Molden, S., Moser, M. B., & Moser, E. I. (2005). Microstructure of a spatial map in the entorhinal cortex. Nature, 436(7052), 801-806. doi.org/10.1038/nature03721
Jacobs, J., Kahana, M. J., Ekstrom, A. D., Mollison, M. V., & Fried, I. (2010). A sense of direction in human entorhinal cortex. PNS, 107(14), 6487-6492. doi.org/10.1073/pnas.0911213107
Jensen, O., Kaiser, J., & Lachaux, J. P. (2007). Human gamma-frequency oscillations associated with attention and memory. Trends Neurosciences, 30(7), 317-324. doi.org/10.1016/j.tins.2007.05.001
Lipman, Fergusson, L., Bonshek, A., Orme-Johnson, D., & Schneider, R. (2021). The Impact of Maharishi Vastu Architecture on Health: A Review of Theory and Research. Medicina, 57.
Lubar, J. F. (1997). Neocortical dynamics: Implications for understanding the role of neurofeedback and related techniques for the enhancement of attention. Applied Psychophysiology and Biofeedback, 22, 111-126. doi.org/10.1023/A: 1026276228832
Moser, E. I., Moser, M. B., & McNaughton, B. L. (2017). Spatial representation in the hippocampal formation: a history. Nature Neuroscience, 20, 1448-1464. doi.org/10.1038/nn.4653
Moser, M. B., & Moser, E. L. (2016). Where Am I? Where Am I Going? Scientific American, 314(1), 26-33. doi.org/10.1038/scientificamerican0116-26
O'Keefe, J., & Dostrovsky, J. (1971). The hippocampus as a spatial map. Preliminary evidence from unit activity in the freely-moving rat. Brain Research, 34(1), 171-175. doi.org/10.1016/0006-8993(71)90358-1
O'Keefe, J., Nadel, L., Keightley, S., & Kill, D. (1975). Fornix lesions selectively abolish place learning in the rat. Experimental Neurology, 48(1), 152-166. doi.org/10.1016/0014-4886(75)90230-7
Petsche, M., Kaplan, S., von Stein, A., & Filz, O. (1997). The possible meaning of the upper and lower alpha frequency ranges for cognitive and creative tasks. International Journal of Psychophysiology, 26, 77-97. doi.org/10.1016/S0167-8760(97)00757-5
Santos-Pata, D., Zucca, R., Low, S. C., & Verschure, P. (2017). Size Matters: How Scaling Affects the Interaction between Grid and Border Cells. Front. Comput. Neurosci., 11, 65. doi.org/10.3389/fncom.2017.00065
Singer, W. (1999). Neuronal synchrony: A versatile code for the definition of relations? Neuron, 24(1), 49-65. doi.org/10.1016/S0896-6273(00)80821-1
Taube, J. S. (1995). Head direction cells recorded in the anterior thalamic nuclei of freely moving rats. Journal of Neuroscience, 15(1), 70-86. doi.org/10.1523/JNEUR0SCI.15-01-00070.1995
Thatcher, R. W., Krause, P. J., & Hrybyk, M. (1986). Cortico-cortical associations and EEG coherence: a two- compartmental model. Electroencephalography and Clinical Neurophysiology, 64(2), 123-143. doi.org/10.1016/0013-4694(86)90107-0
Wang, C. X., Hilburn, I. A., Wu, D. A., Mizuhara, Y., Couste, C. P., Abrahams, J. N. H., Bernstein, S. E., Matani, A., Shimojo, S., & Kirschvink, J. L. (2019). Transduction of the Geomagnetic Field as Evidenced from alpha-Band Activity in the Human Brain. eNeuro, 6(2). doi.org/10.1523/ENEUR0.0483-18.2019
Авторские права
Авторские права на эту статью сохраняются за авторами, а право первой публикации предоставляется журналу.
Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution ( creativecommons.org/licenses/by/4.0/ ).